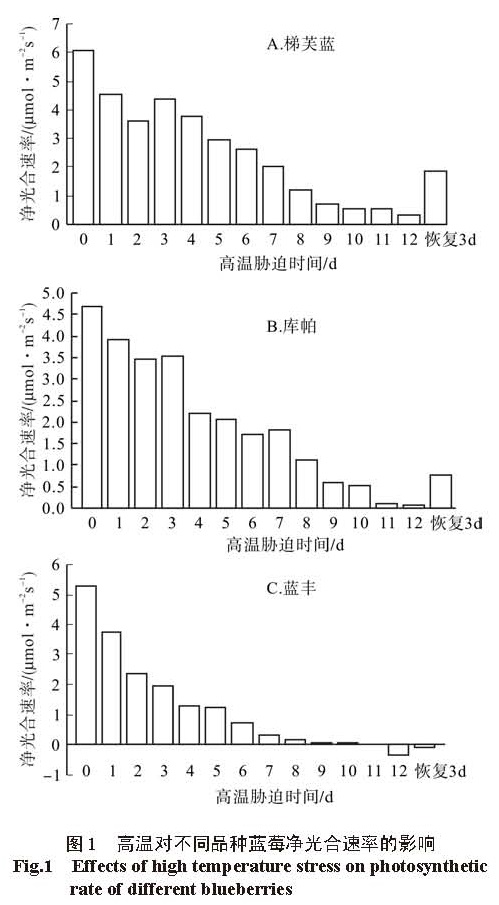
‘库帕’则增加,且‘蓝丰’的增幅大于‘库帕’。高温对 3个蓝莓品种光合速率影响的原因较为复杂,3个品种的蓝莓的光合速率的下降是由气孔限制因素引起的;胁迫后期,光合速率与气孔导度一致下降,胞间 CO2浓度( Ci)呈上升趋势,从这个方面而言,3个蓝莓品种的抗高温能力为‘梯芙蓝’>‘库帕’>‘蓝丰’,这与 3个蓝莓品种大田栽培的表现一致。
3.1 高温对蓝莓光合作用的影响
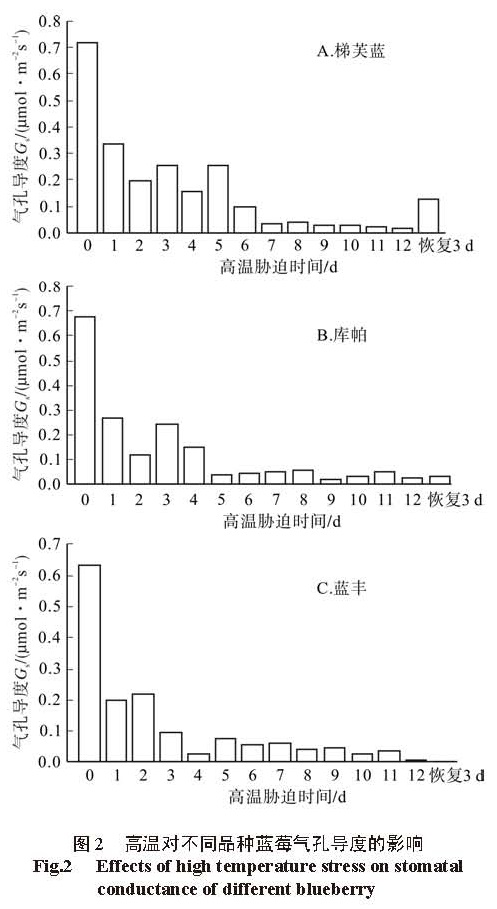
植物的光合作用对环境的变化敏感,高温作为重要的环境因子之一,其对植物光合作用的伤害会出现在受害症状出现之前 [8]。高温对植物的植物光合作用的影响包括气孔限制因素和非气孔限制因素,气孔限制因素强调植物光合作用的下降主要是由于高温胁迫造成气孔导度降低,光合作用的原料 CO2的供应不足导致光合作用下降;非气孔限制因素则认为,高温抑制了光合作用关键酶的活性,叶肉细胞的气体吸收扩散受阻, CO2溶解下降,光合作用的场所稳定性下降,固定 CO2的能力下降等,由此造成光合作用下降 [9]。也有人认为高温对植物光合作用的影响既有气孔限制因素也有非气孔限制因素,是两者共同作用引起的 [10]。本研究表明, 3个品种的蓝莓的净光合速率( Pn)与气孔导度均随着高温胁迫时间的延长表现为降低的趋势,但胁迫前期出现小幅的回升,且两者回升的时间基本一致,说明在高温胁迫前期, 3个品种的蓝莓的光合速率的下降是由气孔限制因素引起的;胁迫后期,光合速率与气孔导度一致下降,胞间 CO2浓度( Ci)呈上升趋势,说明此时光合速率的下降是由非气孔限制因素引起的。由此可见,高温对 3个蓝莓品种光合速率影响的原因较为复杂,尹赜鹏等研究表明,在植物受到外界环境胁迫时,随胁迫时间的增加,光合速率的下降由气孔限制因素转化为非气孔限制因素,与本研究结果一致 [11]。
但不同品种蓝莓的气孔限制因素与非气孔限制因素的时间不同,‘梯芙蓝’在胁迫 1~ 8 d,‘库帕’在胁迫 1~ 5 d,‘蓝丰’在胁迫的 1~ 5 d表现为气孔限制因素,其中蓝丰在胁迫 3~ 5 d是气孔限制因素与非气孔限制因素的综合。气孔限制因素只是对气孔随胁迫时间的延长逐步关闭,使净光合速率下降,此时高温还未对植物内部的生理造成严重影响,等高温解除后,气孔又能恢复正常水平,这从一定程度上表明不同植物耐高温的时间 [12]。从这个方面来说,3个蓝莓品种的抗高温能力为‘梯芙蓝’>‘库帕’>‘蓝丰’,另外高温胁迫解除 3 d后,‘梯芙蓝’的净光合速率恢复程度大于‘库帕’,而‘蓝丰’的光合速率依然几乎为 0,没有恢复。这也进一步验证了‘梯芙蓝’的抗高温能力大于‘库帕’,‘库帕’大于‘蓝丰’,研究结果与这 3个品种的光合实验能相互验证 [13]。
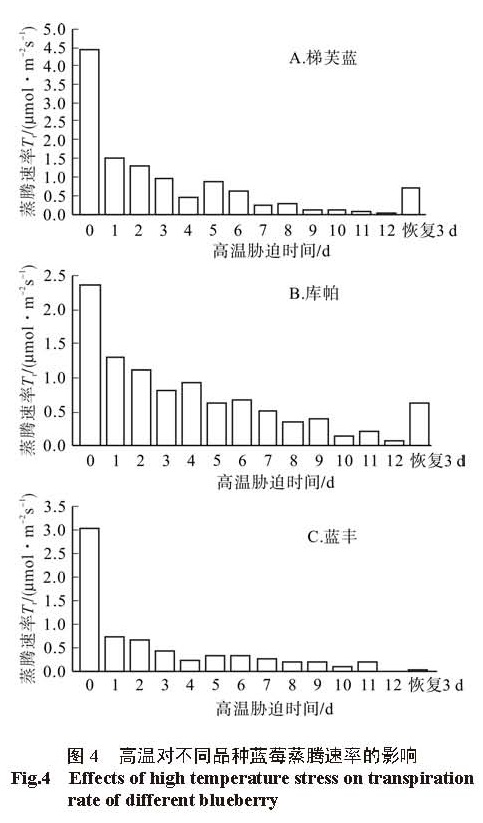
一般认为蒸腾速率与光合作用所产生的能量,土壤中的水分,空气的湿度以及植物自身的生理状况有关 [14]。本研究发现蒸腾速率随着高温胁迫时间的延长而下降,3个蓝莓品种的气孔导度( Tr)与蒸腾速率的下降趋势一致,但气孔导度的下降幅度大于蒸腾速率的下降幅度,这说明蒸腾速率的下降是由气孔导度下降引起的,气孔导度的下降是植物在遇到逆境时,降低自身生理活动,抵御外界胁迫的一种自我保护机制 [15]。
3.2 高温对蓝莓荧光参数的影响
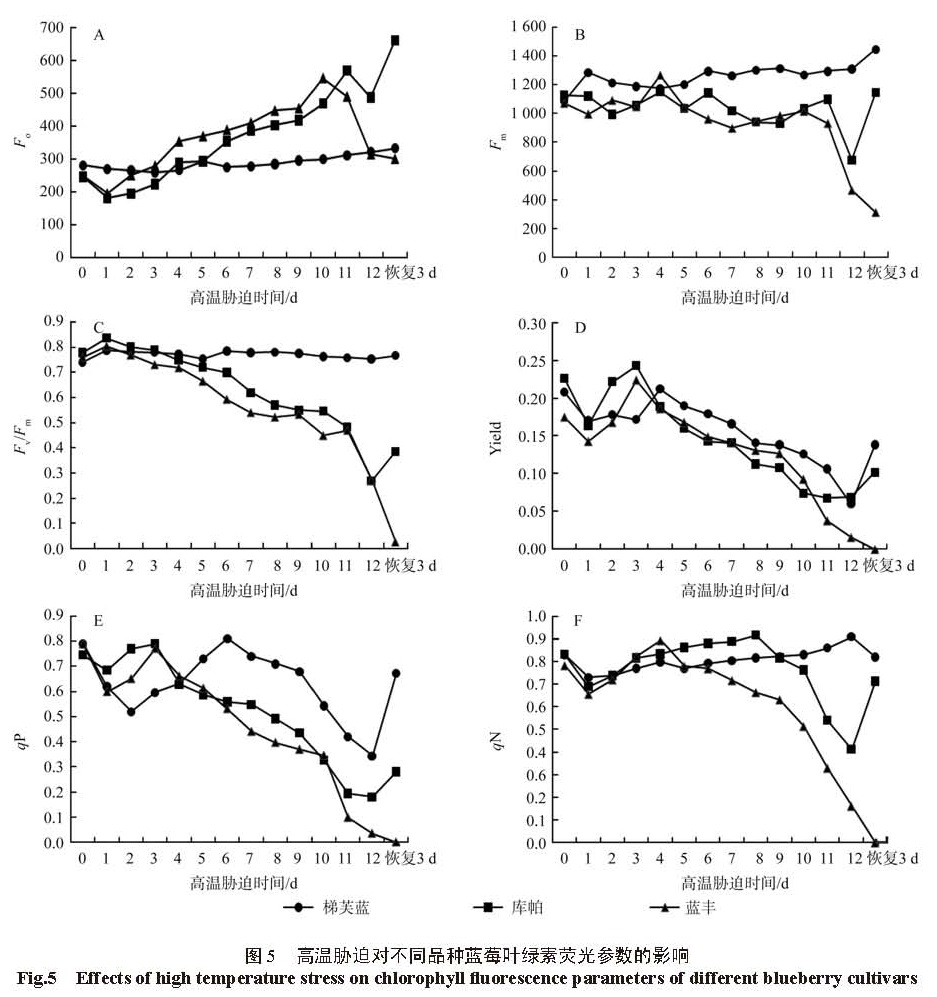
高温不仅影响通过影响气孔的开闭程度,植物体内各种酶的活性来影响植物的光合作用过程,高温也伤害光合反应中心 PSII[16]。光合反应中心 PSII则通过调节反应中心开放程度,光化学量子传递效率等来适应高温的胁迫,保持 CO2同化能力,并通过增加热散耗减轻过剩的能力对自身的伤害 [17]。F0表示在黑暗条件下,反应中心 PSII全部开放的荧光水平,即此条件下,原初电子受体 (QA) 全部氧化,如果反应中心 PSII受到破坏出现可以失活或者不可以的破坏,则 F0就会增加,且增幅越大代表受破坏程度越严重 [5,18]。研究表明在胁迫过程中,3个蓝莓品种在胁迫第 1天,F0都有不同程度的下降,这表明适量高温可促进植物体内的生理活动,随着胁迫时间的延长,梯芙蓝的 F0变化依然不大,呈波浪状略微上升趋势,说明‘梯芙蓝’具有较强的耐高温特性,高温对其反应中心 PSII伤害不大;‘蓝丰’和‘库帕’均随着胁迫时间的延长而增加,且‘蓝丰’的增幅大于‘库帕’,这说明高温对‘蓝丰’的伤害比‘库帕’严重,‘蓝丰’在胁迫 12 d,F0下降,此时‘蓝丰’生理遭受到严重破坏,常温恢复 3 d后,F0回升,这可能是‘蓝丰’在常温环境中进行了部分自我修复;‘库帕’在胁迫第 11 d,F0迅速下降,此时‘库帕’的枝叶已枯黄,生理受到严重伤害,常温恢复 3 d后,F0继续下降,此时植株已死亡。
最大荧光( Fm)是 PSII 反应中心在完全关闭情况下的荧光水平,最大荧光值与传递给 PSII 的电子的量及植物进行光合作用的产物的量呈正相关 [19],它反映了 PSII的电子传递情况, Fv/Fm是最大光化学量子产量,是反映 PSII 光化学效率的指标,高温胁迫下其值下降,下降程度与植物受胁迫程度正相关 [20]。Fm与 Fv/Fm都是判断植物是否受到光抑制的重要标准 [21]。研究结果表明,‘梯芙蓝’在胁迫过程中 Fm与 Fv/Fm变化均不明显,胁迫后期 Fv/Fm略有下降趋势,常温恢复 3 d后,又上升,说明‘梯芙蓝’能承受 43℃的高温;‘库帕’和蓝丰的 Fv/Fm随高温胁迫的时间均下降,‘蓝丰’更敏感,下降幅度更大。这说明‘梯芙蓝’较耐高温,在高温胁迫下仍能保持较高的光化学特性,‘库帕’和蓝丰则因为高温胁迫导致了光抑制或光破坏,其中‘蓝丰’受到破坏的程度更大。本研究还发现,3个蓝莓品种在高温胁迫的第 1天, Fv/Fm均小幅上升。
Yield是 PSII的实际光化学量子产量,即在实际环境中,反应中心部分关闭的条件下,原初电子捕获光能的效率 [5]。也即 PSⅡ反应中心开放的程度与电子传递活性和传递速率呈正相关。研究表明, 3个蓝莓品种在高温胁迫第 1天,Yield就迅速下降,这可能是因为植物长时间在较适宜的环境中生长,突然施加高温胁迫,植物自身迅速关闭部分 PSⅡ反应中心,以抵御高温的伤害 [22]。但从第 3天开始,3个蓝莓品种适宜了高温的环境, Yield增加,经过短暂的上升期后, Yield均大幅下降,说明 3个蓝莓品种均不同程度地受到高温的胁迫,光化学效率降低,有机物积累减少。胁迫恢复 3 d后,‘蓝丰’的 Yield为 0,‘库帕’和‘梯芙蓝’均有回升,且‘梯芙蓝’梯芙蓝的回升幅度大于‘库帕’,说明高温解除后‘梯芙蓝’能较好恢复,‘库帕’受到一定程度的伤害,‘蓝丰’完全死亡。
qP是光化学淬灭,其值反映的是 PSⅡ中的天线色素所吸收的光能用于光化学电子传递的部分,其值越高, PS Ⅱ反应中心开放的比例就越大 [6]。另外,其值越高,还原态 QA重新氧化为 QA的量也越大,电子传递活性就越强 [23]。本研究发现在胁迫初期 3种蓝莓的 qP均显著下降,随后短期上升后下降,这与 Yield对高温的响应一致,原因均可能是因为植物突然受到高温胁迫,迅速关闭部分反应中心以抵御高温的伤害,适应了高温后 Yield开始回升,之后由于高温的伤害, Yield开始下降。但在胁迫第 6天之后,‘梯芙蓝’的 Yield就显著高于‘库帕’和‘蓝丰’,说明‘梯芙蓝’在高温胁迫下能保持较高的电子活性。 qN(非光化学淬灭)系数则反映了 PS Ⅱ反应中心所吸收的光能不能用于光合作用而以热散耗掉的部分能力,当植物吸收过多光能时,热散耗可以消耗部分能量,防止光抑制,可以保护光合系统免受损害 [7]。 3个蓝莓品种的 qN在高温胁迫第 1天均下降,而后上升,这可能是因为启动热散耗来防止光抑制需要时间。从胁迫第 2天开始梯芙蓝的 qN就开始上升一直到胁迫结束才下降,胁迫过程中‘梯芙蓝’可以利用一直增加的热散耗来保护光合系统面受伤害,这也是其 qP较‘库帕’与‘蓝丰’高的原因 [21]。‘库帕’的 qN在胁迫 2~ 9 d上升趋势明显,第 9天以后开始下降,说明‘库帕’在高温胁迫的前 9 d不断增加 qN来增加热散耗,防止光抑制,但胁迫 9 d以后开始下降,这可能是因为生理受到破坏,但结束胁迫恢复常温 3 d后,qN迅速上升,这可能是‘库帕’光合系统自我修复的结果。‘蓝丰’的 qN在胁迫的第 5天即开始下降,胁迫第 9天下降幅度增大,胁迫结束后几乎为 0,说明‘蓝丰’在高温环境第 5天以后已经不能通过增加 qN来增加热散耗,保护自己的光合系统了。通过增加 qN使植物增加热散耗以适应逆境,只是植物被动适应环境的一种方式,这种方式不断增加内损耗,仅能维持生存,但对植物的生长不利且不可持续。
分析 3个蓝莓品种的叶绿素荧光参数对高温胁迫的响应,可以发现植物在突然受到高温胁迫时比较敏感,光合系统 PSⅡ的的开放程度迅速降低,电子活性与传递速率降低,经过短暂的适应后,光合系统 PSⅡ的活性增强,随着高温胁迫时间的增加,光合系统 PSⅡ遭到遭到破坏或者可逆性失活 [4]。另外,蓝莓在逆境中启动自我保护机制,但不同品种蓝莓的自我保护能力不同,因此不同蓝莓的抗高温特性不同,其中‘梯芙蓝’的抗高温特性优于‘库帕’,‘库帕’优于‘蓝丰’,这与高温对光合特性的影响的研究结果一致。同时还发现在高温胁迫下 Yield、qP、qN的敏感性较 F0、Fm、Fv/Fm高。
本研究仅仅是通过测定高温条件下 3个蓝莓品种的部分生理指标,推断不同蓝莓品种的耐热性,而蓝莓的生长除了受温度影响外,还受土壤、水分和肥料等多个因素的影响,因此要筛选出适合高温高湿条件下栽培的蓝莓品种还需要在土壤、肥料等方面开展更多研究,从而综合判定各蓝莓品种的适栽条件。