基金项目:国家自然科学基金项目( 31360473、31260186);新疆维吾尔自治区园艺学重点学科基金项目(201107)
作者简介:李鹏,博士研究生通讯作者:李疆,教授,博士生导师;E-mail:lijiangxj@163.com
作者简介:李鹏,博士研究生通讯作者:李疆,教授,博士生导师;E-mail:lijiangxj@163.com
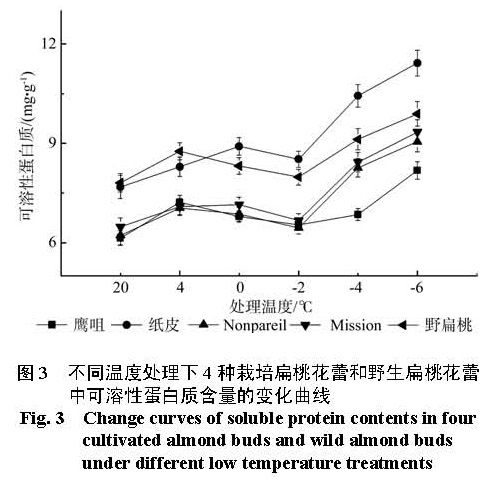
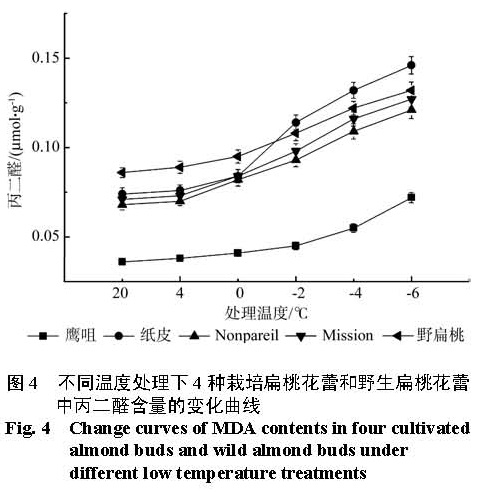
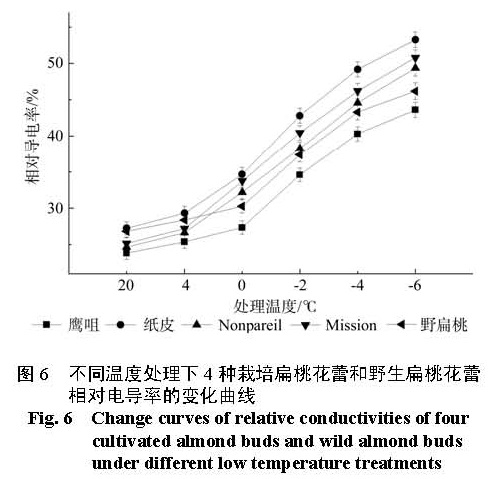
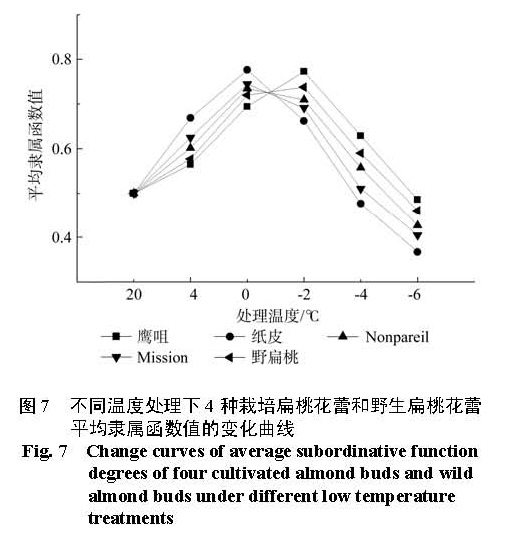
为探明低温下中国栽培扁桃鹰咀、纸皮,国外引进扁桃 Nonpareil、Mission以及野生扁桃花蕾的生理指标变化,比较扁桃花蕾的抗寒性。本实验剪取 4种栽培扁桃和野生扁桃的带花蕾枝条,带回实验室后分别进行 4 ℃、0 ℃、-2 ℃、-4 ℃和 -6 ℃处理,12 h后采摘花蕾,测定花蕾的可溶性糖含量、淀粉含量、可溶性蛋白质含量、 MDA含量、脯氨酸含量和相对电导率变化,并利用隶属函数法比较花蕾的抗寒性。结果 显示低温处理 12 h后,随着处理温度的降低,可溶性糖含量、可溶性蛋白质含量先升高后降低再升高;淀粉含量先降低后升高; MDA含量、相对电导率逐渐升高;脯氨酸含量先升高后降低。利用隶属函数评估法比较发现花蕾的抗寒性强弱顺序是鹰咀>野生扁桃> Nonpareil> Mission>纸皮。
In this research, in order to explore the physiological indexes of buds of the four cultivated almonds (Yingzui and Zhipi originated in China; Nonpareil and Mission introduced from abroad) and wild almond under low temperature, some branches with buds of the four cultivated almonds and wild almond were brought back to the laboratory as experimental materials. And the branches were subsequently processed separately under different low temperatures of 4 ℃ , 0 ℃、-2 ℃ , -4 ℃ and -6 ℃ ; then after 12 hours, the buds were picked off to measure the contents of soluble sugar, starch, soluble protein, MDA and proline as well as the relative conductivity and compare their cold resistance with the method of subordinative function. The results showed that: under different low temperature treatments for 12 hours, the contents of soluble sugar and soluble protein both experienced the change of high-low-high with the temperature declining; the contents of starch went up at first and then fell down; the contents of MDA and the degrees of relative conductivity rose gradually and the contents of proline changed from increasing to decreasing. Thus, the cold resistance degrees of buds of the four cultivated almonds and wild almond exhibited in an order as: Yingzui > wild almond > Nonpareil > Mission > Zhipi.

中南林业科技大学学报
《中南林业科技大学学报》原名《中南林学院学报》,是中南林业科技大学主办的以林为特色的自然科学学术期刊。该刊1981年创刊,2010变更为月刊,月底出版,国内外公开发行。国际刊号为ISSN 1673-923X,国内刊号为CN43-1470/S。该刊是教育部优秀科技期刊,全国优秀高校学报,湖南省一级期刊。是全国中文核心期刊,中国科学引文数据库来源期刊,中国精品科技期刊,中国科技核心期刊。该刊入编了国内所有的期刊数据库。
主要栏目:林学、生态学、生物科学与技术、植物学、园林、木材科学等。
读者对象:本刊适合于农林院校师生以及农林科研院所、农林管理部门和生产单位的科技和管理人员阅读,也适合于与以上学科和专业有关的其他高校师生和科技人员阅读。